|
Кві 04 |
Токсоплазмоз – новый погляд на стару проблемуКатегорія: А.М. Бугаев, доктор ветеринарної медицини про дослідження токсоплазми. Автор: admin |

Автор: А.М. Бугаев, доктор ветеринарної медицини, директор Центру охорони здоровья тварин м.Київ
З сайта http://med-dovidka.com.ua/
 Протягом 17 років (з 1960 по 1977 рік) автор цієї статті брав участь в проведенні експериментальних досліджень по вивченню ендогенних стадій розмноження Anaplasmaovis, Toxoplasma gondii та Besnoitia besnoiti на базі лабораторії токсоплазмозу Інституту зоології АН КазССР. Дослідження проводилися спільно з професором В.Ф.Новинський, а також В.М.Петешевим і У.Д.Вустіной під керівництвом академіка АН КазССР І.Г.Галузо і професора Б.П.Всеволодова. Частина результатів, отриманих в ході наукової роботи, була опублікована в матеріалах, представлених на третьому Міжнародному конгресі протозоологов (1969), а також в книгах “Діагностика токсоплазмозу тварин” (1971) і “Життєвий цикл токсоплазм” під редакцією І.Г.Галузо (1974). На жаль, в книзі “Життєвий цикл токсоплазм” в розділі “Ендогенний цикл розвитку токсоплазм”, написаної мною, крім безстатевої фази розвитку в організмі проміжних господарів були викладені результати наших перших експериментів з вивчення статевої фази розвитку T. gondii, проведених в 1970 році. Як з’ясувалося пізніше, кошенята, яких ми взяли в експеримент, були заражені якимось невідомим нам в той час паразитом, і спори цього збудника були прийняті мною за розділення зиготи токсоплазм (малюнок 1). Після цього було проведено ряд вдалих експериментальних заражень цистами токсоплазм інтактних кошенят і отримані цікаві дані, як по статевій фазі розвитку T. gondii, так і по безстатевому їх розмноженню. Спільно з В.М.Петешевим ми провели унікальні дослідження з вивчення ендогенного розвитку B. besnoiti в організмі великої рогатої худоби. Однак після тієї фатальної помилкової публікації і смерті академіка І.Г. Галузо в 1977 році завдяки старанням “доброзичливців” мої наукові статті довгі роки не публікували. Висловлюю щиру подяку редакції журналу “Нові медичні технології”, особливо головному редактору Д.Є.Афанасьєву, за надану можливість опублікувати справжню роботу і представити ряд особистих архівних матеріалів, які до сих пір не бачили світло. Висловлюю надію, що викладений в статті матеріал, отриманий за допомогою цитологічних, гістологічних та гістохімічних досліджень, буде цікавий і корисний лікарям, а також дослідникам токсоплазмозу і всієї групи токсоплазмідозов. Всі малюнки мікропрепаратів, наведені в статті, – з архіву автора.
Протягом 17 років (з 1960 по 1977 рік) автор цієї статті брав участь в проведенні експериментальних досліджень по вивченню ендогенних стадій розмноження Anaplasmaovis, Toxoplasma gondii та Besnoitia besnoiti на базі лабораторії токсоплазмозу Інституту зоології АН КазССР. Дослідження проводилися спільно з професором В.Ф.Новинський, а також В.М.Петешевим і У.Д.Вустіной під керівництвом академіка АН КазССР І.Г.Галузо і професора Б.П.Всеволодова. Частина результатів, отриманих в ході наукової роботи, була опублікована в матеріалах, представлених на третьому Міжнародному конгресі протозоологов (1969), а також в книгах “Діагностика токсоплазмозу тварин” (1971) і “Життєвий цикл токсоплазм” під редакцією І.Г.Галузо (1974). На жаль, в книзі “Життєвий цикл токсоплазм” в розділі “Ендогенний цикл розвитку токсоплазм”, написаної мною, крім безстатевої фази розвитку в організмі проміжних господарів були викладені результати наших перших експериментів з вивчення статевої фази розвитку T. gondii, проведених в 1970 році. Як з’ясувалося пізніше, кошенята, яких ми взяли в експеримент, були заражені якимось невідомим нам в той час паразитом, і спори цього збудника були прийняті мною за розділення зиготи токсоплазм (малюнок 1). Після цього було проведено ряд вдалих експериментальних заражень цистами токсоплазм інтактних кошенят і отримані цікаві дані, як по статевій фазі розвитку T. gondii, так і по безстатевому їх розмноженню. Спільно з В.М.Петешевим ми провели унікальні дослідження з вивчення ендогенного розвитку B. besnoiti в організмі великої рогатої худоби. Однак після тієї фатальної помилкової публікації і смерті академіка І.Г. Галузо в 1977 році завдяки старанням “доброзичливців” мої наукові статті довгі роки не публікували. Висловлюю щиру подяку редакції журналу “Нові медичні технології”, особливо головному редактору Д.Є.Афанасьєву, за надану можливість опублікувати справжню роботу і представити ряд особистих архівних матеріалів, які до сих пір не бачили світло. Висловлюю надію, що викладений в статті матеріал, отриманий за допомогою цитологічних, гістологічних та гістохімічних досліджень, буде цікавий і корисний лікарям, а також дослідникам токсоплазмозу і всієї групи токсоплазмідозов. Всі малюнки мікропрепаратів, наведені в статті, – з архіву автора.

(Розрив оболонки і вихід спор невідомого паразита (можливо, це пакет яєць цестоди Dipylidium felis) в роздавленої краплі суспензії фекалій кошеня. Зб. 900. 42 42005)
Токсоплазмоз – зоонозное, природно-антропургічне захворювання, що викликається одноклітинними паразитами Toxoplasma gondii, для яких характерна фазність розвитку: в організмі проміжних господарів – фаза облигатно внутрішньоклітинного розвитку (фаза тахізоітів), що закінчується утворенням псевдоцист, і позаклітинна фаза брадізоітів, що завершується утворенням тканинних цист в головному мозку; в організмі Дефінітівних господарів – внутрішньоклітинна, прогамная, внутрішньоепітеліальна фаза, що завершується утворенням гаметоцист, гамет і ооцист; поза організмом – фаза споруляції. Захворювання характеризується унікальними взаємовідносинами паразит – господар і, як наслідок, – різноманітною клінікою.
Проблема токсоплазмозу привернула особливу увагу вчених після того, як був повністю розшифрований життєвий цикл T. gondii і встановлено, що захворювання може бути опортуністичною інфекцією у людини при СНІДі та ряді інших імунодефіцитних станів. Експерти Всесвітньої організації охорони здоров’я в 1972 році включили токсоплазмоз в групу зоонозних захворювань, найбільш небезпечних для здоров’я людини.
Біологія токсоплазм
Однією з біологічних особливостей адаптації T. gondii в організмі проміжного господаря є відсутність у паразита генетично детермінованих, суворо наступних одна за одною стадій розвитку. Будь-яка стадія розвитку токсоплазм (спорозоїт, трофозоіт, ендозоіт, мерозоіт, цістозоіт) може дати початок новому зараженню і нового циклу розвитку в організмі тварини, що є проміжним господарем. Це означає, що будь-яка з перерахованих стадій є інвазивною, причому при будь-якому шляху проникнення токсоплазми в організм тварини і людини. Експериментальне зараження тварин можна здійснити при введенні їм паразитів перорально, підшкірно, внутрішньом’язово, внутрішньочеревно і внутрішньоцеребрально (Галузо І.Г. і співавт., 1974). Нам (Бугаєв А.М., Вустіна У.Д., 1974) вдавалося експериментально заражати білих мишей внутрішньоцеребрально ооцистами слабовірулентного музейного штаму “LTI”. За даними J.P. Dubey і I.K. Frenkel (1972), прогамна фаза розвитку токсоплазм, навіть в кишечнику кішок, не є строго детермінованою. Автори виділили при цьому п’ять структурних типів, які розмножуються асинхронно і можуть переходити один в одного.
Не менш важливою особливістю токсоплазм, що відрізняє їх від інших паразитичних найпростіших класу Sporozoa, є здатність в фазу утворення тканинних цист в головному мозку розмножуватися позаклітинно.
Як показали численні дослідження, проведені в різних країнах світу, у T. gondii існує три незалежних один від одного циклу розвитку. Перший можна спостерігати в організмі дефінітивного господаря при зараженні його ооцистами T. gondii. У цьому циклі беруть участь тільки ссавці – представники сімейства котячих, і зміна господарів тут необов’язкова. Найбільш повно цей цикл вивчили американські дослідники J.P. Dubey і I.K. Frenkel (1972). Згідно з їхніми даними, спорозоїти, що проникли в слизову оболонку тонкого кишківника, що вийшли з ооцист, не розмножуються в її епітелії і не дають прямого початку статевого фази розвитку. На 10-у добу після зараження паразити залишають кишківник, поширюючись в організмі зараженої тварини по кровоносних і лімфатичних судинах. На 18-ту добу токсоплазми, що пройшли безстатеву фазу розвитку в клітинах різних органів, повертаються в покривний епітелій слизової оболонки тонкої кишки. І тільки після цього вони можуть дати початок досить складної безстатевої епітеліальної фази розмноження і розвитку, яка завершується утворенням гаметоцист, гамет і ооцист. Останні разом з калом виділяються неспорулірованими в зовнішнє середовище. Таким чином, весь цикл від ооцисти до ооцисти триває 25-30 діб.
Латентний період при цьому складає 5-20 діб. З результатів досліджень J.P. Dubey і I.K. Frenkel (1972) випливає важливий висновок: для того, щоб у токсоплазм почалася статева фаза розвитку в епітелії кишечника представника котячих, паразити повинні обов’язково пройти фазу вегетативного розвитку в інших клітинах тієї ж або іншої тварини.
Другий цикл (він найбільш повно вивчений багатьма дослідниками, в тому числі і нами) проходить за участю проміжних і дефінітивних господарів. Його тривалість залежить від фази розвитку токсоплазм в організмі проміжних господарів. Якщо відбувається пероральне зараження дефінітивного господаря тахізоітамі, то тривалість циклу від тахізоіта до ооцисти дорівнює 10-15 діб. Якщо ж кішки, особливо кошенята, заражаються брадізоітамі (цістозоітамі), то препатентний період скорочується до 3-5 діб. Препатентний період залежить від того, наскільки “зрілими” або диференційованими є цістозоіти (Пак В.С., 1974). При цьому слід зазначити, що у заражених тахізоітамі і брадізоітамі кошенят поряд з кишковим циклом розмноження і розвитку, що закінчується утворенням ооцист T. gondii, частина паразитів, як і при зараженні ооцистами, проходить кишковий бар’єр, викликаючи генералізований інфікування багатьох внутрішніх органів, яке завершується утворенням тканинних цист в головному мозку і м’язах.
У 1975 році А.М. Бугаєвим і У.Д. Вустіной був проведений експеримент по всередині м’язового зараження 16 кішок у віці від 8 до 20 місяців 2 млн трофозоітов високовірулентні штаму токсоплазм “РН” і 3 кішок у віці 14 місяців цистами слабовірулентнимі штаму “LTI” (не були опубліковані дані). Протягом чотирьох місяців ні в однієї тварини ооцисти в фекаліях виявлені не були. У той же час в місці інокуляції і в паренхіматозних органах кішок паразити виявлялися у великій кількості. Це означає, що статева фаза і препатентний період розвитку токсоплазм залежать від вірулентності, ступеня диференціації паразитів і воротінфіцірованія дефінітивного господаря (кішки).
Третій цикл T. gondii проходить в організмі тварин, які є виключно проміжними господарями. Це більш ніж 300 видів ссавців і птахів, включаючи людину. В їхньому організмі безстатеве розмноження і розвиток паразитів може здійснюватися безперервно за різними схемами: спорозоїт – тахізоіт – брадізоіт, тахізоіт – брадізоіт або ж брадізоіт – тахізоіт – брадізоіт. Підтвердженням наявності таких циклів розвитку у T. gondii є приклади багаторічного (до 35 років!) Підтримки в лабораторних умовах різних штамів токсоплазм на білих мишах, щурах, кроликах і в культурах клітин. При цьому штами можна підтримувати на стадії розвитку тільки тахізоітов, викликаючи гострий токсоплазмоз у мишей і кроликів, або брадізоітов – при роботі зі слабовірулентнимі штамами (Вустіна У.Д., 1974). У природних умовах фази тахізоіта і цисти (брадізоіта) можуть переходити один в одного нескінченно довго і підтримуватися шляхом зараження проміжних господарів ооцистами, які виділяються кішками, а також при поїданні інвазованих органів і тканин заражених тварин. М.Н. Мельник і співавтори (1978) вважають, що для такого циклу розвитку токсоплазм досить тільки аліментарного і конгенітальной механізмів передачі, щоб забезпечити інтенсивну циркуляцію збудника серед тварин, особливо хижаків.
Українські дослідники М.Н. Мельник, Л.К. Коровицький і співавтори (1978), розглядаючи біологію токсоплазм і патогенез токсоплазмозу в організмі проміжного господаря, виділяють дві фази розвитку паразитів – фазу проліферації, що завершується утворенням псевдоцист, і фазу утворення тканинних цист в головному мозку і м’язовій тканині. Автори вважають, що терміни “псевдоцисти” і “циста” мають право на існування,.
У B. besnoiti достовірно встановлено тільки один шлях передачі від тварини до тварини – трансмісивний (Хван М.В., 1968; Bigalke R.D., 1960). З огляду на цикл розвитку у інших видів безноітій, можна припустити, що B. besnoiti в результаті еволюційного перетворення “втратили” свою статеву фазу і перейшли до моноксенному циклу розвитку. Дані В.М. Петешева і А.П. Поломошнова (1974) про виявлення статевої фази розвитку у дикої кішки (Felis libusa), експериментально зараженої матеріалом (цистами B. besnoiti), взятим від спонтанно інфікованих корів, ніхто більше не підтвердив. Треба думати, що, як і в наших перших дослідах, коли кошенята були чистими, так і в експерименті згаданих авторів, кішки були заражені іншими паразитами, оскільки за ними встановилося тверде уявлення як про морфологічних ознаках токсоплазм на певній стадії їх розвитку, так і про інвазійних процесі, який відбувається в організмі людини або тварини.
В останні роки за допомогою методу полімеразної ланцюгової реакції (ПЛР) отримують позитивні результати тестів на токсоплазмоз в пробах крові, слини, сечі, калу, виділених з піхви і очей у тварин з хронічним і латентним перебігом захворювання (неопубліковані дані І.П. Гаврилової – лабораторія молекулярної діагностики при Державному научноконтрольном інституті біотехнологій і штамів мікроорганізмів мапу). Останньому фактору лікарі, по видимому, приділяють недостатньо уваги при розгляді питань патогенезу захворювання та оцінки епідеміології та епізоотології токсоплазмозу.
Таким чином, T. gondii мають дивовижну здатність адаптуватися до організму господаря і постійно “намагаються” перейти від факультатівногетероксенного (двухозяінного) циклу розвитку до моноксенному (однохозяінному). Токсоплазмам, як вказував І.Г. Галузо (1974), “… в загальному життєвому циклі абсолютно не обов’язкова зміна чергування господарів: дефінітивний – проміжний – дефінітивний”. Такої ж думки дотримувався і Д.Н. Засухин (1973), вважаючи, що проходження кишкових стадій розвитку не є облігатним для збереження T. gondii як виду. А якщо врахувати, що у спорідненого токсоплазмам мікроорганізму B. besnoiti взагалі відсутня статева фаза розвитку, то, мабуть, мають рацію були І.Г. Галузо і Д.Н. Засухин, які стверджували, що “такого розвитку ми не знаємо ні у одного загону споровиков”.
Взаємовідносини паразит – господар
Характерною особливістю, яка визначає відносини паразит – господар, а так само і патогенез токсоплазмозу у людини і тварин, є унікальна спо можності T. gondii розмножуватися в багатьох клітинах організму господаря, в тому числі в цитоплазмі клітин ретикулоендотеліальної системи (РЕС) і лейкоцитах (малюнки 2 , 3), а також, в фазу освіти цист, – внеклеточно в тканини головного мозку.
 (Малюнок 2. Колонії тахізоітов в цитоплазмі мезотеліальних клітин брижі білої ми ши, умертвіння через 26 годин після зара ження вірулентним штамом “CDN”. Пластинчастий препарат, окр. за Романовським Гімзою. Ув. 900.)
(Малюнок 2. Колонії тахізоітов в цитоплазмі мезотеліальних клітин брижі білої ми ши, умертвіння через 26 годин після зара ження вірулентним штамом “CDN”. Пластинчастий препарат, окр. за Романовським Гімзою. Ув. 900.)
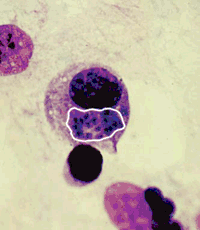 (Малюнок 3. псевдоцисти в моноціт Бриж ки білої миші, умертвіння через 30 го дин після зараження. Пластинчастий препарат, ок. за Романовським Гімзою. Ув. 900.)
(Малюнок 3. псевдоцисти в моноціт Бриж ки білої миші, умертвіння через 30 го дин після зараження. Пластинчастий препарат, ок. за Романовським Гімзою. Ув. 900.)
Оскільки практикуючим медичним працівникам і ветеринарним лікарям, па разітологам і імунологам доводиться мати справу в основному з безстатевою фазою розвитку токсоплазм, ми розглянемо взаємини паразит – господар на примі ре тільки проміжних господарів. Для переконливості наявного в нашому розпорядженні фактичного матеріалу і зроблених нами висновків ми використовували неопубліковані унікальні архівні матеріали Л.А. Бугаевой по цитохімії ендогенного розвитку безноітій в організмі великої рогатої худоби. Це біопсійний матеріал шкіри, який брали у чотирьох спонтанно заражених тварин протягом двох років. Тварини в лабораторію токсоплазмозу Інституту Зоолого гии АН КазССР були доставлені В.М. Петешевим з неблагополучного господарства Балхашского району Талдикурганской області КазРСР в 1975 р і перебували під наглядом до жовтня 1977 р
В організмі проміжних господарів токсоплазми можуть викликати гострий, підгострий і хронічне захворювання. Деякі дослідники виділяють ще й латентний перебіг. При гострому токсоплазмозі відносини паразит – господар добре простежуються при зараженні вірулентними штамами токсоплазм білих мишей і молодих тварин інших видів. А.М. Бу гаїв і В.Ф. Новинський (1969-1974) викликали експериментально гострий перебіг за болевания у білих мишей і цуценят шляхом внутрішньом’язового і внутрішньочеревно введення вирулентного штаму токсо плазм “СDN”. Досвідчені тварини заражені лись трофозоітов, отриманими з екс судата у білих мишей, на яких підтримувався штам. У перші дві доби після зараження токсоплазми посилено розмножувалися в цитоплазмі мезотеліальних кле струм серозних оболонок черевної порожнини. У пластинчастих препаратах брижі білих мишей, забарвлених за Романовським Гімзою, чітко видно як розмноження токсоплазм, так і зміна внаслідок цього морфології інвазованих клітин. Бінарне і множинне розподіл токсоплазм ми докладно описали в ті зісах доповіді до третього Міжнародного конгресу протозоологов, що відбувся в 1969 р, і в книзі “Життєвий цикл токсоплазм”, виданої за редакцією І.Г. Галузо (1974). Тут ми останавлива емся тільки на взаєминах токсоплазм з організмом – господарем.
 (Малюнок 5. Гістологічний зріз печінки білої миші, умертвіння на 5е добу після зараження. мікронекрози з великою кількістю токсоплазм навколо тих, хто гине гепатоцитів. Ок. залізним гематоксиліном. Ув. 900.)
(Малюнок 5. Гістологічний зріз печінки білої миші, умертвіння на 5е добу після зараження. мікронекрози з великою кількістю токсоплазм навколо тих, хто гине гепатоцитів. Ок. залізним гематоксиліном. Ув. 900.)
 (Малюнок 4. Пластинчастий препарат брижі білої миші, умертвіння через 24 години після зараження. Колонія тахізоітов з хоро шо вираженою паразітофорной вакуолью. Ядро инвазированной клітини лизировать, а поруч розташовані клітини гіпертрофується вани і мають по 23 ядерця. Ок. по Рома новскомуГімзе. Ув . 900.)
(Малюнок 4. Пластинчастий препарат брижі білої миші, умертвіння через 24 години після зараження. Колонія тахізоітов з хоро шо вираженою паразітофорной вакуолью. Ядро инвазированной клітини лизировать, а поруч розташовані клітини гіпертрофується вани і мають по 23 ядерця. Ок. по Рома новскомуГімзе. Ув . 900.)
Протягом перших 24-48 годин після за ражения паразити викликають помітну ги пертрофію інвазованих клітин. При чому, як показали цитохимические дослі нання (Бугаєва Л.А., 1974), не тільки в інвазованих мезотеліальних клітинах, а й у клітинах запального инфильтра та, що утворюється навколо токсоплазм, де спостерігається підвищений метаболізм (малюнок 4). Цитохімічних це подтвержда ється підвищенням кількості РНК і загального білка як в цитоплазмі, так і в ядерцях клітин. Однак у міру збільшення числа паразитів в інвазованих клітинах піро нінофілія цитоплазми зменшується. Слабшає і забарвлення по Мейзі для виявлення загального білка. У інвазованих клітинах виникають виражені дистрофічні зміни у вигляді вакуолизации або зернистої дистрофії. При оптимальному для токсоплазм перебігу патологічного процесу оболонка клітини руйнується і збудники, вільно залишаючи її, потрапляють в черевну порожнину. Потім, очевидно, частина з них знову впроваджується в кліть ки мезотелия і моноцити і активно в них розмножується, після чого з потоком крові розноситься по всьому організму, викликаючи паразитемию. На 4-5е добу після зара ження паразити вже виявляються у внутрішніх органах. У гіршому для збудників випадку, інвазовані клітини під впливом безпосередньо паразитів і їх токсинів піддаються некробі ОЗУ і некрозу (малюнок 5). Разом з клітина ми макроорганізму дистрофічний про токсіліном. Ув. 900.
 (Малюнок 6. Гістологічний зріз печінки цуценя, зараженого токсоплазмами штаму “CDN”. псевдоцисти в клітці Купфера, ядро клітини півмісяцевою форми і відсунуто па тхне до периферії. Ок. залізним гематоксиліном. Ув. 900)
(Малюнок 6. Гістологічний зріз печінки цуценя, зараженого токсоплазмами штаму “CDN”. псевдоцисти в клітці Купфера, ядро клітини півмісяцевою форми і відсунуто па тхне до периферії. Ок. залізним гематоксиліном. Ув. 900)
 (Малюнок 7. Сформовані цисти в корі головного мозку цуценя, знищену на 30е добу після зараження. у брадізоітов позитивна реакція на амилопектин. Біла речовина навколо цист отечное, періцелюлярние простору нейронів різко розширені. Ок. по Макманус. Ув. 900.)
(Малюнок 7. Сформовані цисти в корі головного мозку цуценя, знищену на 30е добу після зараження. у брадізоітов позитивна реакція на амилопектин. Біла речовина навколо цист отечное, періцелюлярние простору нейронів різко розширені. Ок. по Макманус. Ув. 900.)
процес (пикноз і рексіс) охоплює і самих збудників. Коли тварини гинуть внаслідок гострого перебігу захворювання, псевдоцисти і цисти ще не встигають утворитися. Однак у цуценят, які пережили гостру фазу токсоплазмозу (в нашому експерименті у 5 з 20), на 17е добу після зараження в клітинах РЕС виявлялися псевдоцисти (рисунок 6), а на 30-добу в головному мозку були виявлені цисти токсоплазм (малюнок 7). Цитохімічні дані (Бугаєва Л.А., 1974) свіедетельствовалі про те, що перехід фази тахізоіта в фазу брадізоіта супроводжується зменшенням вмісту в цитоплазмі паразитів РНК і загального білка. Що ж стосується полісахаридів (амілопектину), то при цьому спостерігається поступове їх на накопичені в цитоплазмі збудника. Вна чалі в псевдоцисти полісахариди видно у вигляді дрібної зернистості, а в брадізоітах вони зливаються у великі конгломерати і відсувають ядро до заднього кінця клітини. Така ж тенденція пізніше була подтверж Дена в дослідженнях Т.В. Бейер (1978).
Проблема токсоплазмозу є складною і багатогранною. Протягом більше 90 років вона привертає увагу вчених різних спеціальностей. При цьому вивчення питань токсоплазмозной інвазії у людини і тварин присвячено велику кількість робіт, а історія вивчення проблеми повне важливих общебиологических відкриттів, які докорінно змінили уявлення про поділ паразитичних найпростіших і систематичне положення не тільки токсоплазм і всієї групи токсоплазмоподобних паразитів, але і представників загону Cocidiida .
До 1970 р були практично вивчені основні питання, пов’язані з біологією токсоплазм, їх циркуляцією в природі, ендогенних циклом розвитку, епідеміологією і епізоотологією, а також патогенезом, клінікою, діагностикою та навіть лікуванням. Однак з відкриттям статевої фази розвитку у T. gondii і токсоплазмоподобних організмів спостерігається спроба “переосмислити” всі ці питання з позиції кокцідіозних, а вірніше ейміріозних понять. Це в якійсь мірі ввело в оману, як спільнота клінічної медицини, так і ветеринарних лікарів. У науковій літературі щодо токсоплазм і токсоплазмоподобних організмів з’явилася незрозуміла термінологія: “справжні і несправжні кокцидии”, “цистоутворюючої кокцидии”, “нееймерідние кокцидии”, “тканинні кокцидии”. Окремі “ентузіасти” в своїх фантазіях дійшли до порівняння складних взаємин токсоплазм і організму хазяїна з “троянським конем”. Автор сподівається, що справжня публікація дозволить залучити до дискусії лікарів різних спеціальностей, які внесуть свій вагомий внесок у вирішення не тільки проблеми токсоплазмозу, а й усієї чітко відокремленої групи токсоплазмоподобних організмів. Тоді все прикметники за типом “справжні” і “несправжні” кокцидии зникнуть з термінології токсоплазмід.
У процесі відносин паразит-господар при токсоплазмозі та інших токсоплазмідозах інтерес представляє факт освіти збудником паразітофорних вакуолей. Проведені нами дослідження показали, що ці структури формуються навколо паразитів, як розташованих усередині клітин, так і знаходяться вільно в рідинах організму хазяїна, наприклад в серозному ексудаті (рисунок 8). Подібні паразітофорние вакуолі ми виявляли і в ядрах клітин РЕМ при безноітіозе великої рогатої худоби.
Детальне вивчення методом світлової мікроскопії паразітофорних вакуолей різних внутрішньоклітинних паразитичних найпростіших показало, що вони складаються з дрібнозернистої або гомогенної маси, оточеній тонкою мембраною. При електронно-мікроскопічному дослідженні (Sheffild, Hammont, 1967; Colley, 1968) паразітофорная вакуоль представляється електронносветлой, заповненої аморфно-гранульованим вмістом. Вона оточена двошарової мембраною. Створюється враження, з одного боку, що вакуоль разом з мембраною захищає паразита від потужних окислювальних процесів, що відбуваються в клітині при впровадженні в неї збудника, з іншого – що клітина таким чином захищається від впливу патогена. Автори вважають, що паразітофорная вакуоль утворюється в результаті гідролізу частини цитоплазми клітини під впливом виділяються збудниками ферментів. Мабуть, ці ферменти розщеплюють білки та інші мікронутрієнти цитоплазми клітини до більш низькомолекулярних сполук, які добре засвоюються паразитами. Таким чином, паразітофорная вакуоль служить як би “комори”, в якій знаходяться готові запаси поживних речовин, що забезпечують зростання і розвиток патогенного мікроорганізму. Виявлення паразітофорной вакуолі навколо збудників, що знаходяться в ексудаті альвеол легенів, свідчить про ферметатівном механізмі її утворення, а не по типу “троянського коня”, як це уявляє собі Т.В. Бейер (2004).
Феномен гідролізу навколишнього паразита тканини ми виявили не тільки в фазу гострого перебігу токсоплазмозу, а й при утворенні тканинних цист токсоплазм і токсоплазмоподобних паразитів. У токсоплазм це добре проглядається при утворенні цист в тканини головного мозку тварин. При переході захворювання з гострої фази в хронічну паразити після освіти в клітинах РЕС псевдоцист залишають внутрішні органи і поселяються в головному мозку, скелетних м’язах і серце.

Рисунок 9
(Малюнок 9. Різні фази розвитку позаклітинних шизонтів в білій речовині кори головного мозку (показано однією стрілкою) і вільна, позаклітинна колонія токсоплазм, навколо якої оболонка цисти відсутня (показано двома стрілками). Ок. По Макманус (кольоровий знімок), і залізним гематоксиліном (чорно-білий знімок). Ув. 900.)

Рисунок 10
(Малюнок. 10. Гістологічні зрізи кори головного мозку білого миші зараженої внутрішньом’язово штамом “LTI” і умертвіння на 9е добу після зараження. Гомогенізовані тканину (гідроліз) білої речовини навколо позаклітинних брадізоітов. Ок. По Мак Мануу. Ув. 900. )

Рисунок 11
(Малюнок. 11. Різні стадії розвитку тканинних цист токсоплазм в стовбурової частини головного мозку білого миші, зараженої штамом “LTI” і умертвіння на 20е добу після зараження. Гістологічний зріз. Ок. По Макманус. Ув. 900.)
Проведені нами експерименти на білих мишах, собак і кішок показали, що в головному мозку цисти токсоплазм утворюються внеклеточно і тільки в білій речовині. Формування цист починається з позаклітинного розмноження токсоплазм шляхом шизогонії або бінарного розподілу (малюнок 9). Як і при внутрішньоклітинному розвитку в фазу тахізоітов, токсоплазми, що потрапили в головний мозок, спочатку викликають гомогенизацию (гідроліз) білої речовини (рисунок 10) і лише потім утворюється оболонка цисти. Цитохімічні дослідження Л.А. Бугаевой (1974) показали, що гомогенізує частина білої речовини дає різко позитивну ШИК-реакцію, позитивну реакцію на РНК, містить велику кількість негістонових білків, а по ВанГізон забарвлюється в світло-зелений колір, т. Е. Дає негативну реакцію. З цього випливає важливий висновок, що гомогенезірованная тканина не є результатом фібриноїдного набухання нервових волокон або їх некрозу, що підтверджується фарбуванням цих ділянок в Світлосиній колір за методом Сельє (некротизованих тканина при фарбуванні цим методом набуває різні відтінки оранжевого та рожевого кольорів). Треба думати, що і в білій речовині головного мозку ферменти токсоплазм викликають не некроз і не фибриноидное набухання, а гідроліз. Таким чином, паразити готують живильне середовище для подальшого свого розвитку, але вже в складі колонії.
Після утворення значного числа паразитів навколо їх колонії починає формуватися оболонка (рисунок 11). Дослідники одностайні в думці, що токсоплазми в цисте продовжують розмножуватися і розвиватися. Добре вивчена морфологія і цитохімія цистної стадій розвитку токсоплазм. Так, С.М. Пак (1974), вивчаючи морфологію брадізоітов, звернув увагу на те, що розташування ядер в них різне. Він прийшов до висновку, що токсоплазми з центральним розташуванням ядер складають основну масу молодих цист, тоді як в більш зрілих цистах велика частина представлена паразитами з кінцевим розташуванням ядер. Л.А. Бугаєва (1974), що дослідила морфологію і цитохімія брадізоітов, показала, що в їх цитоплазмі, в порівнянні з тахізоітамі, міститься більше амілопектину, а місцезнаходження ядер пов’язано з накопиченням полісахаридів. У брадізоітах з центральним розташуванням ядер амилопектин у вигляді зерен розподілений по всій цитоплазмі, а при ядрі, зміщеному до полюса клітини, він зібраний в одну велику гранулу, яка займає центральне положення. Таким чином, очевидно, що оболонка цисти утворюється в результаті складного взаємини колонії T. gondii з організмом господаря. На думку Д.Н. Засухіна (1974), цисти не є спочиває стадією – в них безперервно відбувається розмноження паразитів, що веде до збільшення розміру освіти. Тоді виникає питання про харчування декількох тисяч мікроорганізмів усередині цисти. Відповідь на нього ми знаходимо в роботі англійської патоморфолога Beverley (1969), який, вивчаючи ультраструктуру стінки цист токсоплазм, виділених з тканини головного мозку білих мишей, встановив, що вона складається з оболонки товщиною 150-200 ангстрем, яка містить безліч канальців, що забезпечують інтенсивний обмін між організмом господаря і колонією паразитів. Автор вважає, що освіта цист не залежить від імунного стану макроорганізму, а є нормальною фазою життєвого циклу токсоплазм.
Для розуміння механізму утворення тканинних цист у токсоплазмоподобних паразитичних найпростіших, зокрема у токсоплазм, ми розглянули деталі цього процесу у B. besnoiti (рисунок 12). У B. besnoiti найбільш чітко виражені всі морфологічні і біологічні особливості ендогенного розвитку токсоплазмід.
При розмноженні і розвитку B. besnoiti в організмі тварин, так само, як і у токсоплазм в організмі проміжних господарів, чітко простежується дві фази – проліферації, що закінчується утворенням внутріядерних псевдоцист, і формування позаклітинних цист. У підгостру і хронічну фази перебігу захворювання безноітіі, як і токсоплазми, утворюють псевдоцисти. Відмінність полягає лише в тому, що псевдоцисти токсоплазм виявляються в цітомлазме клітин РЕМ і лейкоцитах, а при безноітіозе їх можна бачити всередині ядер різних клітин, в тому числі і клітин РЕС. У фазу освіти внутріядерних псевдоцист безноітіі, як і токсоплазми, проникнувши в каріоплазма клітини, не тільки формують паразітофорную вакуоль, а й якимось чином гидролизуют цитоплазму инвазированной клітини і навколишнє її тканину (рисунок 13). При цьому гідролізу піддаються всі клітини і міжклітинний речовина, що оточують вогнище (ядро) інвазії, в тому числі клітинні елементи запального інфільтрату і багатошаровий плоский епітелій (рисунок 14). Гидролізірованний тканину дає різко позитивну піронінофіліі і реакцію на загальні білки. У ній виявляється велика кількість полісахаридів (рисунок 15). В осередку процесу безноітіі розмножуються, а потім, в більшості випадків, виходять з нього в гомогенізований масу (рисунок 16). Нерідко можна спостерігати картину виходу паразитів в гомогенізований тканину з двох або трьох і більше ядер псевдоцист (рисунок 17). Вийшовши з них, безноітіі продовжують розмножуватися вже внеклеточно (як і токсоплами!), Що підтверджується збільшенням інтенсивності забарвлення ядер мікроорганізмів на ДНК і посиленням піронінофіліі їх цитоплазми (рисунок 18). Після утворення великої позаклітинної колонії можна побачити формування товстої тришарової оболонки. При цьому створюється враження, що в колонії існує “спеціалізація” паразитів. Організми, які розташовані по периферії колонії, містять у своїй цитоплазмі більшу кількість амілопектину в порівнянні зі збудниками, що знаходяться в центрі колонії. У останніх менш виражена піронінофіліі. Характерно, що у паразитів, розташованих
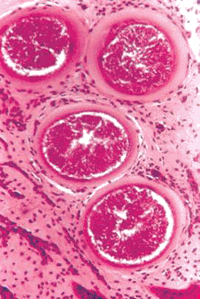 (Малюнок 12. Добре сформовані трьох шарові цисти B. besnoiti в дермі спонтанно зараженої корови при хронічному перебігу захворювання. Гістологічний зріз. Ок. гематоксілінеозіном. Ув. 130.)
(Малюнок 12. Добре сформовані трьох шарові цисти B. besnoiti в дермі спонтанно зараженої корови при хронічному перебігу захворювання. Гістологічний зріз. Ок. гематоксілінеозіном. Ув. 130.)
 (Малюнок 13. Гістологічний зріз біопатов шкіри 11месячного бичка, спонтанно що заразився B. besnoiti. Широка зона гомогенізований тканини навколо ядер інвазії безноітіямі (псевдоцист). Ок. залізним гематоксиліном. Ув. 900.)
(Малюнок 13. Гістологічний зріз біопатов шкіри 11месячного бичка, спонтанно що заразився B. besnoiti. Широка зона гомогенізований тканини навколо ядер інвазії безноітіямі (псевдоцист). Ок. залізним гематоксиліном. Ув. 900.)
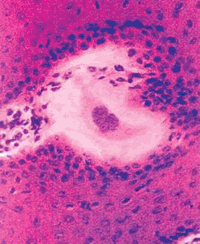 (Малюнок 14. Гістологічний зріз багатошарового плоского епітелію біопатов шкіри бичка. Видно зона гомогенізації клітин росткового, шипуватий і зернистого шарів епідермісу. Ок. залізним гематоксиліном. Ув. 280.)
(Малюнок 14. Гістологічний зріз багатошарового плоского епітелію біопатов шкіри бичка. Видно зона гомогенізації клітин росткового, шипуватий і зернистого шарів епідермісу. Ок. залізним гематоксиліном. Ув. 280.)
на периферії колонії, під впливом неуточнених факторів цитоплазма руйнується, а гранули полісахаридів йдуть на побудову тонкого, внутрішнього шару цистної стінки. Долю ядер таких мікроорганізмів простежити дуже важко. Мабуть, вони гідролізуються під впливом ферментів живих особин.
Середній шар цистної оболонки безноітій повністю формується з гомогенізований маси тканин організму господаря. Цей товстий шар, можливо, грає роль своєрідної “паразітофорной вакуолі”, оскільки містить велику кількість піронінофільних матеріалу, загального білка і полісахаридів (рисунок 19). Третій, зовнішній, шар цисти, також утворюється з гомогенізований маси. Уявлення про те, що оболонка цисти формується з сполучної тканини, склалося, очевидно тому, що в окремих випадках можна бачити, як пучки сполучної тканини піддаються гідролізу і перетворенню в зовнішній шар цистної оболонки (рисунок 20). Важливо підкреслити, що тканинні цисти за своїм внутрішнім змістом неоднорідні. Поряд зі звичайними цистами нерідко в хронічну фазу хвороби всередині сегментованих цист зустрічаються мікроцисти з щільно упакованим зернистим вмістом, забарвлюється в темно-синій колір (рисунок 21). Роль їх в життєвому циклі B. besnoiti поки незрозуміла.
Такий найчастіших ознак спосіб утворення тканинних цист у B. besnoiti. Однак трапляються випадки, коли оболонка инвазионного ядра не рвуться, а в міру розмноження і розвитку паразитів всередині каріоплазми реорганізується і стає внутрішнім шаром товстої цистної оболонки колонії (рисунок 22). При цьому цистної стінка колонії безноітій має не три, а чотири шари.
Аналогічним шляхом відбувається утворення і тканинних цист у саркоспорідій в серці і скелетних м’язах великої рогатої худоби, який є для цих паразитів проміжним господарем. І в цьому випадку після фази проліферації, яку вони проходять в цитоплазмі клітин РЕМ, збудник формує в скелетних м’язах і серце тканинні цисти, освіту яких йде двома шляхами. В одному випадку, і повідомимо частіше, цисти утворюються в
 (Малюнок 15. Гістологічний зріз біопатов дерми корови. ШІКположітельная реакція гомогенізований маси і цитоплазми брадізоітов. Ок. по Макманус. Ув. 350.)
(Малюнок 15. Гістологічний зріз біопатов дерми корови. ШІКположітельная реакція гомогенізований маси і цитоплазми брадізоітов. Ок. по Макманус. Ув. 350.)
 (Малюнок 16. Гістологічний зріз біопатов дерми бичка. Вихід безноітій з внутрішньоядерній псевдоцисти в гомогенізований тканину. Ок. залізним гематоксиліном. Ув. 300.)
(Малюнок 16. Гістологічний зріз біопатов дерми бичка. Вихід безноітій з внутрішньоядерній псевдоцисти в гомогенізований тканину. Ок. залізним гематоксиліном. Ув. 300.)
 (Малюнок 17. Гістологічний зріз біопатов дерми бичка. Вихід в гомогенізований тканину безноітій з трьох ядер псевдоцист. Підгострий перебіг захворювання. Ок. гематоксилін-еозином. Ув. 300.)
(Малюнок 17. Гістологічний зріз біопатов дерми бичка. Вихід в гомогенізований тканину безноітій з трьох ядер псевдоцист. Підгострий перебіг захворювання. Ок. гематоксилін-еозином. Ув. 300.)
 (Малюнок 18. Гістологічний зріз біопатов дерми бичка. ДНК у внутріядерних безноітіях. Реакція Фельгена. Ув. 300.)
(Малюнок 18. Гістологічний зріз біопатов дерми бичка. ДНК у внутріядерних безноітіях. Реакція Фельгена. Ув. 300.)
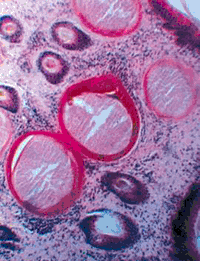 (Малюнок 19. Гістологічний зріз біопатов дерми бичка. РНК в середньому шарі стінки цисти. Реакція Браше. Ув. 200.)
(Малюнок 19. Гістологічний зріз біопатов дерми бичка. РНК в середньому шарі стінки цисти. Реакція Браше. Ув. 200.)
 (Малюнок 20. Гістологічний зріз біопатов дерми бичка. Формування молодої тканинної цисти. Видно гомогенізація окремих волокон сполучної тканини. Ок. по Макманус. Ув. 300.)
(Малюнок 20. Гістологічний зріз біопатов дерми бичка. Формування молодої тканинної цисти. Видно гомогенізація окремих волокон сполучної тканини. Ок. по Макманус. Ув. 300.)
саркоплазме м’язових волокон. Так само як у токсоплазм і безноітій, проникли в саркоплазму м’язових волокон паразити починають розмножуватися і розвиватися. Але до цього вони гідролізують (гомогенизируют) саркоплазму м’язового волокна, а потім, у міру освіти численної колонії паразитів, навколо неї формується оболонка цисти (рисунок 23). Тут ми бачимо утворення оболонки цисти вже навколо колонії паразитів, в якій бере участь гомогенізує саркоплазма. Сарколеммой м’язового волокна ніякої участі в освіті цистної стінки не приймає. Поряд з цим, можна спостерігати утворення цист в міокарді не в одному, а декількох м’язових волокнах – в таких випадках гидролизации піддається три або п’ять з них (малюнок 24). І тут освіту тканинних цист йде за таким же принципом: спочатку паразити викликають гідроліз м’язових волокон з утворенням колонії, а потім, гидролизуя групи м’язових волокон, формують з гомогенної маси оболонку цисти (рисунок 25). При цьому добре помітний активний край цисти, в якому паразити розмножуються, гидролизуя м’язові волокна, і пасивний, де, очевидно, знаходяться гаметоцити. Неактивний край оточений добре сформованої оболонкою. З боку активного краю оболонка цисти відсутня.
 (Малюнок 21. Гістологічний зріз біопатов дерми корови. Темного кольору мікроцисти всередині звичайної тканинної цисти (показано стрілкою). Ок. Залізним гематоксиліном. Ув. 300.)
(Малюнок 21. Гістологічний зріз біопатов дерми корови. Темного кольору мікроцисти всередині звичайної тканинної цисти (показано стрілкою). Ок. Залізним гематоксиліном. Ув. 300.)
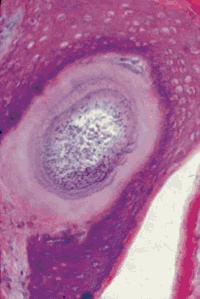 (Малюнок 22. Гістологічний зріз біопатов епідермісу бичка. Ядерна оболонка не розривається, а стає внутрішнім шаром тканинної цисти B. besnoiti. Ок. залізним гематоксиліном. Ув. 300.)
(Малюнок 22. Гістологічний зріз біопатов епідермісу бичка. Ядерна оболонка не розривається, а стає внутрішнім шаром тканинної цисти B. besnoiti. Ок. залізним гематоксиліном. Ув. 300.)
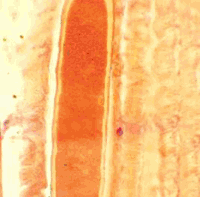 (Малюнок 23. Гістологічний зріз скелетного м’яза, отриманий автором в 1981 р від зебу на Гаванському м’ясокомбінаті, Республіка Куба. Добре сформована циста Sarcocistis sp. в гидролізірованний м’язовому волокні. Видно, що сарколеммой м’язового волокна ніякої участі в освіті оболонки цисти не приймає. Ок. гематоксілінеозіном. Ув. 250.)
(Малюнок 23. Гістологічний зріз скелетного м’яза, отриманий автором в 1981 р від зебу на Гаванському м’ясокомбінаті, Республіка Куба. Добре сформована циста Sarcocistis sp. в гидролізірованний м’язовому волокні. Видно, що сарколеммой м’язового волокна ніякої участі в освіті оболонки цисти не приймає. Ок. гематоксілінеозіном. Ув. 250.)
Обговорення
Численні дослідження в різних країнах світу показали, що токсоплазми дивним чином можуть адаптуватися до організму хазяїна. У даного збудника відсутні строго детерміновані фази розвитку, як це має місце у багатьох паразитичних найпростіших класу Sporozoa. При цьому необов’язковою є зміна проміжних і дефінітивного господарів для підтримки існування виду в природі. Токсоплазми входять в невелику групу паразитичних найпростіших, які можуть розмножуватися в клітинах РЕС і лейкоцитах. Строго внутрішньоклітинна локалізація токсоплазм спостерігається тільки в фазу проліферації (фазу тахізоіта). Якщо навіть розглядати формування цист токсоплазм в саркоплазме м’язових волокон, то і тут процес відбувається за єдиним механізмом, ідентичному такого в тканинах головного мозку. Після проникнення паразита в саркоплазму м’язового волокна спочатку відбувається її гомогенізація (гідроліз) і розподіл паразитів, і тільки після утворення колонії токсоплазм формується оболонка цисти. Цей же принцип освіти тканинних цист ми спостерігаємо і у родинних токсоплазмам паразитів – саркоцист. Тут так само, як і у токсоплазм і безноітій, спочатку гідролізується навколишнє тканина, готується “поживна і будівельна платформа”, а потім вже формується оболонка цисти.
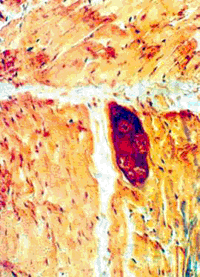 (Малюнок 24. Гістологічний зріз міокарда зебу. Початкова стадія позаклітинної цисти Sarcocistis sp. Видно діляться метроціти в гомогенізований тканини. Ок. гематоксілінеозіном. Ув. 250.)
(Малюнок 24. Гістологічний зріз міокарда зебу. Початкова стадія позаклітинної цисти Sarcocistis sp. Видно діляться метроціти в гомогенізований тканини. Ок. гематоксілінеозіном. Ув. 250.)
 (Малюнок 25. Гістологічний зріз міокарда зебу. Формування цисти Sarcocistis sp. в пучку м’язових волокон. Ок. гематоксілінеозіном. Ув. 120.)
(Малюнок 25. Гістологічний зріз міокарда зебу. Формування цисти Sarcocistis sp. в пучку м’язових волокон. Ок. гематоксілінеозіном. Ув. 120.)
Якщо проводити аналогію з кровепаразітов, які розмножуються в еритроцитах і лейкоцитах, то токсоплазми в стадію проліферації – це паразити паренхіматозних органів і переважно клітин РЕМ, а в фазу освіти тканинних цист – паразити головного мозку, скелетних м’язів і серця. Д.Н. Засухин і Г.Т. Акиншина (1973) описали цисти в сполучної тканини. При цьому автори пов’язували дану локалізацію з тим, що головний мозок, м’язи і сполучна тканина імунологічно малоактивні. Нам же видається, що такого роду тропность токсоплазмід пов’язана з високим вмістом полісахаридів в зазначених тканинах, а в фазу розвитку цист їм необхідна велика кількість амілопектину.
Таким чином, очевидно, що не тільки токсоплазми, а й інші споріднені з ними збудники (безноітіі і саркоцистами) з суворо внутрішньоклітинних паразитів в фазу інтенсивної проліферації перетворюються в колоніально розвиваються паразитів, які можуть розмножуватися як внутрішньоклітинно, так і внеклеточно зі своєю “спеціалізацією” в межах колонії (B. besnoiti). Їх внутрішньоклітинна облігатно закінчується в фазу гострого перебігу захворювання або в фазу інтенсивної проліферації і формування псевдоцист. У токсоплазмід відзначається тенденція переходу від строго гетерогенного циклу розвитку, який має місце у Sarcocystis, до факультативно гетерогенного (у T. gondii) і моноксенному (у B. Besnoiti).
Про місце Toxoplasma gondii в системі найпростіших
Після відкриття статевої фази розвитку у токсоплазм і деяких схожих з ними паразитичних найпростіших з’явився ряд доказів, що спонукають ще раз повернутися до питання про місце токсоплазмід в системі найпростіших.
Не всі токсоплазміди мають статеву фазу розвитку, яка протікає по типу кокцидий. Навіть ті токсоплазміди, у яких є фаза статевого розвитку, що проходить по типу ізоспорід, мають дуже складну фазу безстатевого розвитку, яка ніяк не вписується в схему “справжніх” або “нецістообразующіх” кокцидий. Дані J.P. Dubey і J.K. Frenkel (1972) про наявність у I. felis і I. rivolta фази проліферації в організмі гризунів підтверджують таксономічну зв’язок ізоспорід і токсоплазмід. Однак їх фази проліферації в організмі проміжних господарів настільки різняться, що це може бути аргументацією на користь відмінності цих паразитів на рівні родин. За ендогенного розвитку і розмноження в організмі проміжних господарів токсоплазміди більш схожі з паразитами роду Theileria, ніж з ізоспори.
У T. gondii і B. besnoiti відсутні строго детерміновані стадії розвитку. У токсоплазм, навіть в кишечнику дефінітивного господарів, відзначається асинхронність розмноження – немає послідовності в розвитку окремих форм, як це має місце у кишкових кокцидий (Dubey, Frenkel, 1972).
У жодного представника “справжніх” кокцидий немає чергування облигатно внутрішньоклітинного і внеклеточно-колоніального розвитку, як у токсоплазм, безноітій і саркоцист.
У жодного представника “справжніх” кокцидий немає такої фази освіти тканинних цист, як у токсоплазм, безноітій і саркоцист. Фактично в більшості випадків цисти зазначених видів паразитів є результатом позаклітинного, колоніального розвитку брадізоітов.
У токсоплазм і саркоцист статева фаза розвитку в організмі дефінітивного господаря строго залежить від стадій розвитку паразитів в організмі проміжного господаря.
Всі стадії розвитку токсоплазм (за винятком гаметоцитов, гамет і неспорулірованних ооцист) і безноітій інвазивних при будь-якому методі введення паразитів.
Для T. gondii необов’язкова зміна дефінітивного господаря проміжним.
В організмі проміжного господаря можлива безперервна (нескінченна) проліферація T. gondii і B. besnoiti.
B. besnoiti притаманний трансмісивний шлях передачі при відсутності статевої фази розвитку за типом еймереід або ізоспорід.
У токсоплазмід має місце тенденція переходу від облигатно двухозяінного циклу (Sarcocystis і деяких видів Besnoitia) до факультативно облигатному 30 5/2005 У ФОКУСІ 5/2005 31 В ФОКУСІ (T. gondii), аж до моноксенного у B. besnoiti.
З урахуванням перерахованих десяти основних особливостей розвитку токсоплазмід, по видимому, слід погодитися з пропозицією акад. І.Г. Галузо (1974) про виділення їх в самостійний загін підкласу Coccidiomorpha c незначними змінами в нижчих таксонах.
Систематика токсоплазмід в даний час могла б виглядати наступним чином:
Клас Sporozoa, Leuckart, 1879
Підклас Coccidiomorpha, Dofllein, 1901
Загін Coccidiida, Labbe, 1899
Загін Toxoplasmida, Biocca, 1957, Galuso, 1970
Сімейство Toxoplasmidae, Dubey, Fren kel, 1970
Рід Toxoplasma, Nicolle, Manceaux, 1908
Рід Sarcocystis, Ray Lankester, 1882
Рід Besnoitia, Marotel, 1912 Рід Frenkelia, Biocca, 1968
Сімейство Isosporidae, Schneider, 1881
Рід Isospora, Wenyony, 1923.